Get Complete Project Material File(s) Now! »
Secondary and tertiary structure
The structure of the human soluble BAFF consists of two layered antiparallel strands that form a typical jellyroll-like sandwich, like other members of the TNF ligand family (Liu, Xu et al. 2002; Oren, Li et al. 2002). This structure contains 12 strands called: A (aa146-151), A’’’ (aa158-160), A’’ (aa 163-165), A’ (aa 168-174), B’ (aa 178-181), B (184-187), C (aa191-201), D (aa 208-215), E (aa 226-235), F (aa 245-253), G (aa 258-262) and H (aa 270-283) that are organized in two sheets (Karpusas, Cachero et al. 2002; Liu, Xu et al. 2002) (Figure 6). The first anti-parallel sheet forming the jellyroll comprises strands A’ , A, H, C and F and the second, strands B’, B, G, D and E. Homologies between BAFF and some members of the TNF-L family (TNF- TRAIL occur in strands C, D, F, G and H which constitute the core of the jellyroll fold, whereas the loop regions AA’, CD, DE, EF and GH are the most divergent regions among the TNF family ligands (Liu, Xu et al. 2002; Oren, Li et al. 2002). Soluble BAFF presents characteristic of other TNF-ligand family members: its structure in loops CD and EF; the AA’ loop has an insertion of two short strands, A’’’ and A’’, that form a hairpin motif called the “Elbow region”. The AA’ loop, contains a large insert between strands D and E: the “FLAP region” which is unique to soluble BAFF.
Quaternary structure
Like all TNF-ligands (Smith and Baglioni 1987), the biological form of BAFF is a trimer. The interface that forms this trimer mainly consists of layered aromatic residues including the F194, Y196 and Y246 monomers. Two hydrophobic interactions are involved in this BAFF-trimer formation, mediated on one hand by the Q144 from each monomer (hydrogen bond) and on the other hand by the three last residues L282,284,285 from the C-terminus of three monomers (Liu, Xu et al. 2002). The unique FLAP region (DE loop) of BAFF allows trimer-to-trimer interaction leading to a virus-like assembly of the soluble trimers (Liu, Xu et al. 2002). This structure contains 20 trimers associated with each other by hydrogen bonds and hydrophobic bonds involving 4 residues: Y192, K252, E254 and H218. H218 from the FLAP region seems decisive for the formation of oligomers. Indeed, when histidine in position 218 is replaced by an alanine, BAFF cannot oligomerize at pH 7 (Cachero, Schwartz et al. 2006). At pH 6.0, BAFF exists only in a trimeric form. At pH 6.5, the ratio oligomers/trimers is 1:2 and 1:1 at pH 7.0. At pH 7.4, only the oligomeric form is present. The oligomeric form can also induce the proliferation of B cells in vitro with the same efficiency as the trimeric forms. These structural forms were detected in the supernatants of several cell lines, demonstrating their existence (Cachero, Schwartz et al. 2006). However, their physiological role has not yet been clearly demonstrated.
Glycosylation
BAFF has two potential sites for N-glycosylation formed by two asparagines at position 124 and 242. According to Schneider et al., the complete form of BAFF is N-glycosylated on N124 but not on N242 (Schneider, MacKay et al. 1999). After treatment with N-glycanase F, the molecular weight of BAFF decreases. The absence of glycosylation on N242 could be due to the secondary structure of the protein because this residue is present at the beginning of the strand F. The cleaved form of the soluble BAFF should not be N-glycosylated because the cleavage site is downstream N124. However, another team showed that the cleaved form of BAFF was glycosylated on N242 after expression of this soluble form in Pichia pastoris (Diao, Ye et al. 2007). The molecular weight of soluble BAFF was then found to have increased from 17 kDa to 20 kDa.
BAFF-producing cells
BAFF is mainly expressed by mononuclear cells from blood, spleen and lymph nodes, although low expression exists in the placenta, the thymus and the heart (Moore, Belvedere et al. 1999; Mukhopadhyay, Ni et al. 1999; Schneider, MacKay et al. 1999; Shu, Hu et al. 1999; Alsaleh, Messer et al. 2007; Langat, Wheaton et al. 2008).
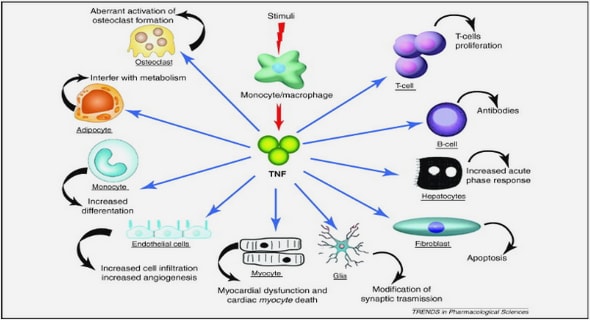
The main innate immune cells that express BAFF are monocytes, macrophages, neutrophils and follicular dendritic cells (DC) (Hase, Kanno et al. 2004). Activated T and B cells also produce BAFF (Huard, Arlettaz et al. 2004; Kern, Cornuel et al. 2004; Daridon, Devauchelle et al. 2007). Other cell types also express BAFF (Mackay, Silveira et al. 2007): stromal cells from the bone marrow (Schaumann, Tuischer et al. 2007), (Ohata, Zvaifler et al. 2005), astrocytes (Krumbholz, Theil et al. 2005), and EC (Daridon, Pers et al. 2006). BAFF was also found in the synovium of patients with rheumatoid arthritis (RA) (Rochas, Hillion et al. 2009).
BAFF-induced production
Myeloid cells such as monocytes, macrophages and monocyte-derived DC release BAFF after IFN- , IFN- and CD40 ligand (CD40L) stimulations (Litinskiy, Nardelli et al. 2002). In these cells, membrane-bound BAFF can be cleaved by a furin convertase to produce a soluble form (Nardelli, Belvedere et al. 2001). However, neutrophils do not express BAFF on their surface and show a special mechanism for BAFF secretion upon stimulation with G-CSF (granulocyte-colony stimulating factor) and IFN- (Scapini, Nardelli et al. 2003). So, whereas BAFF is cleaved at the membrane in other cell types of cells; in neutrophils, it is cleaved intracellularly. Additionally, macrophages, DC and neutrophils synthesize BAFF after IFN- and lypopolysaccharide (LPS) stimulation through the production of reactive oxygens (Moon, Lee et al. 2006). On the contrary, IL-4 inhibits the expression of BAFF in monocytes (Nardelli, Belvedere et al. 2001; Scapini, Nardelli et al. 2003). TGF- upregulates BAFF expression by macrophages. In mouse macrophages, TGF-has been shown to increase BAFF expression through the TGF- signalling pathway where Smad3 and Smad4 promoted BAFF promoter activity. In the same study, IFN- stimulation further increased TGF- -induced BAFF expression through the phosphorylation of CREB, and involved the PKA/CREB pathway in the IFN- induced BAFF expression (Kim, Jeon et al. 2008).
In human intestinal EC, IFN- induces the production of both soluble and membrane bound BAFF through the JAK/STAT signalling pathway and through the binding of phosphorylated STAT-1 to the BAFF promoter (Woo, Im et al. 2013). SOCS3 plays an important role in the induction of BAFF by gut EC because of the IFN- stimulation. Upregulation of SOCS3 blocks the JAK/STAT pathway signalling which in turn suppresses the production of BAFF by IFN- stimulation (Do, Choi et al. 2013).
Macrophage and Dendritic cells
BAFF expression is detected on the surface of human DC and macrophages along with its secretion as a soluble form into culture supernatants. IFN , IFN , IL-10, CD40L, LPS, and peptidoglycan can induce BAFF in these models (Nardelli, Belvedere et al. 2001). Both cells found in the splenic MZ can produce and secrete BAFF (Balazs, Martin et al. 2002). However, the information regarding BAFF production by murine myeloid cell types is more limited and controversial. TLR agonists such as LPS and CpG oligodeoxynucleotides do not induce BAFF secretion by murine DC (Boule, Broughton et al. 2004). However, in another study, the LPS-induced surface expression of BAFF on DC could be observed (Diaz-de-Durana, Mantchev et al. 2006). BAFF induces DC activation and maturation. BAFF activates DC in order to secrete inflammatory cytokines like IL-6, IL-1 , TNF- and to induce the proliferation of naïve CD4+ T cell and their differentiation into effector CD4 T cells so as to modulate the immune response (Chang, Mihalcik et al. 2008). Unlike APRIL, BAFF secreted by these two cells is more effective to induce the costimulation for B cell proliferation through a BCR-dependent pathway (Craxton, Draves et al. 2005). BAFF from macrophages and DC regulates B cell function by enhancing the proliferation, the antibody secretion and by inducing Ig class-switching. In mice, BAFF affects the maturation of follicular DC and, as in BAFF-deficient mice, the mature follicular dendritic network is affected (Rahman, Rao et al. 2003). BAFF also causes dendritic cells to produce various inflammatory cytokines like IL-6, TNF- . During this process, BAFF can induce the proliferation of naïve CD4+ T cells and regulate the differentiation of CD4+ T cells into CD4+ Th1 cells in a DC-mediated manner. And finally, BAFF also elevates the activation and maturation of DC and plays an indirect role in modulating the adaptive immune system (Chang, Mihalcik et al. 2008).
BAFF as a therapeutic target
The importance of BAFF in the mouse model of autoimmunity, the implications of BAFF in the survival of lymphoma cells, and the increased BAFF levels in various diseases have propelled the role of BAFF as a therapeutic target. Several strategies have been developed to block BAFF. Selective inhibition of BAFF is achieved with either soluble BAFF-R or with antibodies to BAFF (Moisini and Davidson 2009).
Selective BAFF blockers prevent BAFF from interacting with its receptors, leaving APRIL free to interact with TACI and BCMA. A clinical program led by Human Genome Sciences in partnership with GlaxoSmithKline has led to the development of a fully human BAFF-specific monoclonal antibody (belimumab; Lymphostat-B) (Baker, Edwards et al. 2003). A fusion protein consisting of human Ig Fc and of the extracellular BR3 domain (Briobacept, for BAFF-R-Ig) has also been developed. Non-selective BAFF blockers abolish the interactions of both BAFF and APRIL with all their receptors. To date, there is one drug in this class (Seshasayee, Valdez et al. 2003). It is based on human Ig Fc fused to the extracellular TACI domain (Atacicept, TACI-Ig). Atacicept has completed its phase I clinical trial and is now being tested in phase II and III clinical trials in the treatment of SLE, RA and relapsing multiple sclerosis. It yields good results with a good safety profile in the treatment of SLE and RA (Carbonatto, Yu et al. 2008). In a recent study, lupus nephritis patients treated with Atacicept showed an unexpected decrease in serum IgG levels, severe proteinuria and increased rates of infection. These results brought the trials to an end (Ginzler, Wax et al. 2012).
The various secreted form of BAFF, caused by translational modifications and heterogeneity, make it difficult to analyze their concentrations in patients. This has led to ongoing controversies because in some groups of patients the serum concentration of BAFF remains within normal range. Hence, monitoring the BAFF levels before and after treatment remains a thorny issue (Mariette, Roux et al. 2003; Collins, Gavin et al. 2006). Additionally, the glycosylation is also problematic because it seems to alter the epitope recognition by anti-BAFF Abs.
History of regulatory B cells
In 1968, Morris et al, first suggested that the suppressive function of B cells was mainly restricted to their ability to produce ‘inhibitory’ Abs (Morris A et al 1968). This finding was then followed by reports linking B cell’s suppressive effect to the induction of tolerance and differentiation of suppressor T cells (L’Age-Stehr, Teichmann et al. 1980; Shimamura, Habu et al. 1984). In experimental autoimmune encephalomyelitis (EAE) model, mice lacking mature B cells were unable to recover from the disease whereas normal mice could. The poor recovery potential of these B cell-deficient mice proves undeniably that B cells play a role in the suppression of EAE (Wolf, Dittel et al. 1996).
Table of contents :
CONTENTS
FIGURES INDEX
ABBREVIATIONS
PREAMBLE
I. INTRODUCTION
1. BAFF
1.1. BAFF forms: from the gene to the protein
1.1.1. Genetics of BAFF
1.1.1.1. BAFF gene an overview
1.1.1.2. Promoter and transcription factors for BAFF
1.1.1.3. Polymorphism
1.1.2. Variants of BAFF
1.1.2.1. BAFF
1.1.2.2. BAFF
1.1.2.3. 4BAFF
1.1.2.3.1. Genetics
1.1.2.3.2. Protein localization and expression
1.1.2.3.3. Splicing regulation of 4BAFF
1.1.3. BAFF protein
1.1.3.1. Introduction
1.1.3.2. Primary structure
1.1.3.3. Secondary and tertiary structure
1.1.3.4. Quaternary structure
1.1.4. Glycosylation
1.2. BAFF-producing cells
1.3. BAFF-induced production
1.4. BAFF Receptors
1.4.1. BAFF-R
1.4.2. TACI
1.4.3. BCMA
1.5. Functions of BAFF
1.5.1. B cells
1.5.2. T cells
1.5.3. Macrophage and Dendritic cells
1.6. BAFF as a therapeutic target
2. Regulatory B cell
2.1. History of regulatory B cells
2.2. Development and activation of regulatory B cells
2.2.1. CD40 signalling
2.2.2. BCR engagement
2.2.3. Toll-like receptors
2.2.4. CD80 and CD86
2.2.5. Other signals inducing regulatory properties of B cells
2.2.6. Phenotype of regulatory B cells
2.3. BAFF and regulatory B cells
2.4. Function of regulatory B cells . How do they work ?
3. Toll-like receptors
3.1. Introduction
3.2. Structure
3.3. Expression and Ligands
3.3.1. Bacterial PAMPs recognized by TLRs
3.3.2. Viral PAMPs recognized by TLRs
3.3.3. Fungal and protozoal PAMPs recognized by TLRs
3.3.4. Endogenous ligands recognized by TLRs
3.4. Signaling pathways
3.5. TLRs and B cells
3.6. Role of TLRs in autoimmunity
4. Primary Sjögren’s Syndrome
4.1. Introduction
4.2. Epidemiology of the disease
4.3. Clinical symptoms and diagnosis
4.3.1. Sicca syndrome
4.3.2. Extraglandular manifestations
4.3.3. Classification criteria
4.4. Immunopathology of the disease
4.4.1. Introduction
4.4.2. Epithelial cells regulate autoimmune epithelitis
4.4.3. Role of T cells in SS
4.4.4. Role of B cells in SS
4.4.5. Role of BAFF in SS
II. ARTICLES
1. SPECIFIC FORMS OF BAFF FAVOR BAFF RECEPTOR-MEDIATED EPITHELIAL CELL SURVIVAL
2. DELTA 4 BAFF IS A TRANSCRIPTION FACTOR ENHANCING THE PRODUCTION OF BAFF AND CONTROLLING REGULATORY B CELL FUNCTIONS
3. TLR9 EXPRESSED ON PLASMA MEMBRANE ACTS AS A NEGATIVE REGULATOR OF HUMAN B CELL RESPONSE
III. GENERAL DISCUSSION
IV. REFERENCES.
GET THE COMPLETE PROJECT