Get Complete Project Material File(s) Now! »
Aims
The aims of this study were to investigate the direct and indirect interactions of cobalt, palladium, platinum and vanadium with human neutrophils in vitro, leading to either hyper-reactivity or under-reactivity of these cells, both of which have adverse health implications. With respect to the former, the pro-oxidative, pro-inflammatory potential of the metals was investigated by measuring their effects (vanadium only) on the generation of the highly toxic reactive oxidant, hydroxyl radical, by activated neutrophils, as well as effects (all of the metals) on the activation and translocation to the nucleus of the cytosolic transcription factor, NF-κB. In the case of the latter, two strategies were used to investigate the indirect interactions of the metals with neutrophils. These were to characterize the effects of the metals on; i) the chemotactic/Ca2+-mobilizing functions of three key chemoattractants in innate host defences viz C5a, and IL-8; and ii) the interactions of the metals with pneumolysin, a toxin produced by Streptococcus pneumoniae, which also triggers innate, protective inflammatory reactions which prevent colonization with this microbial pathogen. S. pneumoniae is the most commonly encountered bacterial pathogen in communityacquired pneumonia, with particularly high mortality rates in the very young, the elderly, and those infected with HIV-1. In this setting, pneumolysin has been used as a prototype microbial activator of innate host defences.
Neutrophils
All cellular elements of the blood arise from haematopoietic stem cells in the bone marrow. These pluripotent cells divide into two more specialized cells, a common lymphoid progenitor and a myeloid progenitor cell. The lymphoid progenitor gives rise to the T- and B-lymphocytes, the myeloid progenitor to basophils, eosinophils, monocytes and neutrophils. Basophils, eosinophils and neutrophils are known collectively as polymorphonuclear leukocytes. Neutrophils have a 14-day development period in the bone marrow and stay temporarily in a storage pool before being released into the blood. There they spend 12 – 14 hours in transit from a circulating pool into a marginating pool where they are in contact with the vessel walls. Thereafter, in the absence of any infection, neutrophils enter reticuloendothelial organs, such as the liver, or even return to the bone marrow to undergo apoptosis (Brown et al, 2006). Following penetration of the mechanical barriers of the host, neutrophils are the first line of defence against bacterial- and fungal infection. However, activated neutrophils can cause extensive harm to host cells and neutrophil-mediated tissue injury may contribute significantly to the pathogenesis of numerous diseases (Ayub & Hallett, 2004). Recently, it has been proposed that an inappropriate activation and positioning of neutrophils within the microvasculature contributes to multiple organ dysfunction and failure in sepsis (Brown et al, 2006).
Calcium homeostasis in activated neutrophils
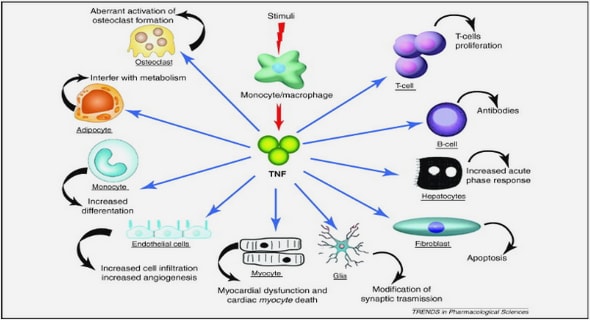
In unstimulated neutrophils cytosolic free Ca2+ is present at very low levels (≈ 100nM). After receptor-mediated activation, there is an abrupt and short-lived increase in cytosolic free Ca2+. This increase in the cytosolic Ca2+ concentration peaks at 10 – 20 seconds, lasts for several minutes and is a prerequisite for the initiation of pro-inflammatory activities of neutrophils. These Ca2+-dependent functions include activation of β2-integrins and adhesion to vascular endothelium, activation of NADPH oxidase and subsequently superoxide production, degranulation, activation of phospholipase A2 and activation of proinflammatory, cytosolic nuclear transcription factors, such as NF-κB. Nuclear transcription factors in turn activate the genes encoding the inflammatory cytokines IL-8 and TNF-α. The Ca2+ may originate exclusively from the intracellular stores, or from both intracellular and extracellular reservoirs and is dependent on the type of receptor- mediated stimulus (Anderson et al, 2000; Tintinger et al, 2005). Intracellular Ca2+ is stored at different sites within the neutrophil. One store is located peripherally under the plasma membrane and seems to be involved in the activation of β2-integrins.
The other site is located in the perinuclear space and is mobilized by chemoattractants, like FMLP, C5a, leukotriene B4, PAF and chemokines via leukocyte membrane receptors, which belong to the G-protein-coupled family of receptors. Binding to these receptors results in the activation of phospholipase C, which, through hydrolysis of phosphatidylinositol, produces inositol-1,4,5- triphosphate (IP3). IP3 interacts with Ca2+ mobilizing receptors on the intracellular storage vesicles, which in turn result in a rapid 5 to 10 fold increase in the cytosolic calcium concentration. Neutrophil apoptosis As neutrophils have the potential to inflict harm to host tissue, it is important that their activity is tightly regulated, especially in inflammation when large numbers of activated neutrophils may accumulate within one organ. These mechanisms are referred to as ‘programmed cell death’ or ‘apoptosis’. Apoptotic neutrophils fragment to form ‘apoptotic bodies’, which can be phagocytosed by macrophages. Prior to this, signalling shutdown may limit the function of the neutrophil. Apoptotic neutrophils are non-functional; they are unable to move by chemotaxis, generate a respiratory burst or degranulate, and there is a clear down-regulation of cell surface receptors, preventing them from transducing signals. The rate of apoptosis may also be accelerated, as well as delayed. Delayed neutrophil apoptosis correlates with severity of clinical sepsis and multiple organ dysfunction (Keel et al, 1997; MatuteBello et al, 1997). Experimentally, bacterial products and pro-inflammatory cytokines delay apoptosis (Colotta et al, 1992).
Interleukin-8 (IL-8)
Neutrophils are capable of generating many different cytokines, chemokines, growth factors and other proteins in vitro and in vivo. The production of individual cytokines by neutrophils is influenced to a great extent by the stimulatory conditions. The chemokine, IL-8, seems to play a critically important role in inflammatory processes. Several studies have identified an association of IL-8 with various acute and chronic inflammatory conditions including sepsis, psoriasis, rheumatoid arthritis, gout, severe trauma, pulmonary fibrosis, asthma, emphysema, pneumonia and adult respiratory distress syndrome (Hoch et al, 1996).
When Pseudomonas aeruginosa was used as a stimulus, it was observed that the bacterium and its products induced IL-8 expression in airway epithelial cells and the recruitment of neutrophils into the airways (Oishi, et al 1994). IL-8 induces activation of G-protein coupled receptors which results in the activation of phospholipase C, which catalyzes the hydrolysis of membrane phosphoinositides to yield diacylglycerol (DAG) and IP3, which in turn mobilizes the intracellular Ca2+, as mentioned above (Rahman, 2000). Significantly elevated levels of IL-8 and myeloperoxidase have been found in sputum of toluene diisocyanate-asthma and dust-asthma patients (Jung & Park 1999). Several studies have shown the potential role of IL-8 in haematopoiesis and trafficking of haematopoietic stem cells. Systemic administration of IL-8 induces rapid mobilization of progenitor cells from the bone marrow (Van Eeden & Terashima 2000; Fibbe et al, 2000). IL-8, as well as GM-CSF and TNF-α are also implicated in the regulation of neutrophil oxidative burst by modulating the activity of the NADPH oxidase through a priming (sensitizing) phenomenon. These cytokines induce a very weak oxidative response by neutrophils but strongly enhance neutrophil release of ROS on exposure to a second stimulus (Gougerot-Pocidalo et al, 2002).
Table of Contents :
- Declaration
- Summary
- Samevatting
- Publications
- Acknowledgments
- Table of Contents
- List of Figures
- List of Tables
- Abbreviations
- Chapter 1: Literature Review
- 1.1 Metals
- 1.1.1 Platinum
- 1.1.2 Palladium
- 1.1.3 Cobalt
- 1.1.4 Vanadium
- 1.2 Neutrophils
- 1.2.1 Neutrophil activation
- 1.2.1.1 Neutrophil recruitment to inflammatory sites
- 1.2.1.2 Neutrophil phagocytosis and degranulation
- 1.2.1.3 Respiratory burst and the NADPH oxidase system
- 1.2.1.4 Calcium homeostasis in activated neutrophils
- 1.2.2 Neutrophil apoptosis
- 1.2.3 Interleukin-8 (IL-8)
- 1.2.1 Neutrophil activation
- 1.3 C5a
- 1.3.1 Introduction
- 1.3.2 C5a as anaphylatoxin
- 1.3.3 C5a as chemoattractant
- 1.3.4 C5a actions on neutrophils, monocytes and macrophages
- 1.4 Pneumolysin
- 1.4.1 Introduction
- 1.4.2 Origin of pneumolysin
- 1.4.3 Structure of pneumolysin
- 1.4.4 Biological effects of pneumolysin
- 1.5 Nuclear Factor-Kappa B
- 1.5.1 Introduction
- 1.5.2 NF-κB proteins
- 1.5.3 The IκB proteins
- 1.5.4 Activation of NF-κB
- 1.5.4.1 Activating signals
- 1.5.4.2 IκB-kinase (IKK)
- 1.5.4.3 Degradation of IκB
- 1.5.4.4 Receptors
- 1.5.4.5 Transcription of NK-κB mediated genes
- 1.5.5 NF-κB and disease
- 1.5.6 NF-κB and apoptosis
- 1.5.7 NF-κB in neutrophils
- 1.1 Metals
- Chapter 2: Vanadium Promotes Hydroxyl Radical Formation by Activated Human Neutrophils
- 2.1 Introduction
- 2.2 Materials and Methods
- 2.2.1 Chemicals and reagents
- 2.2.2 Neutrophils
- 2.2.3 Superoxide anion production
- 2.2.4 Electron spin resonance (ESR) spectroscopy
- 2.2.5 Spectrofluorimetric detection of hydroxyl radical
- 2.2.6 Myeloperoxidase (MPO) activity
- 2.2.7 Cellular ATP levels
- 2.2.8 Expression and statistical analysis of results
- 2.3 Results
- 2.3.1 Hydroxyl radical generation in cell-free systems
- 2.3.2 Superoxide production by activated neutrophils
- 2.3.3 Hydroxyl radical production by activated neutrophils
- 2.3.4 Activity of myeloperoxidase
- 2.3.5 Cellular ATP levels
- 2.4 Discussion
- Chapter 3: Activation of Nuclear Factor-Kappa B
- (NF-κB) in Human Neutrophils
- 3.1 Introduction
- 3.2 Materials and Methods
- 3.2.1 Chemicals and reagents
- 3.2.2 Neutrophils
- 3.2.3 NK-κB activation
- 3.2.4 Interleukin
- 3.2.4.1 Production of IL
- 3.2.4.2 Immunological reactivity of IL
- 3.2.5 Phosphorylated proteins
- 3.2.6 Expression and statistical analysis of results
- 3.3 Results
- 3.3.1 NF-κB activation
- 3.3.1.1 NF-κB activation in pneumolysin-treated neutrophils
- 3.3.1.2 Effects of the metals on NF-κB activation
- 3.3.2 IL
- 3.3.2.1 Production of IL
- 3.3.2.2 Immunological reactivity of IL
- 3.3.1 NF-κB activation
- 3.3.3 Phosphorylated proteins
- 3.3.4 Cellular ATP levels
- 3.4 Discussion
- Chapter 4: Palladium Attenuates the Proinflammatory
- Interactions of C5a, Interleukin-8 and Pneumolysin
- with Human Neutrophils
- 4.1 Introduction
- 4.2 Materials and Methods
- 4.2.1 Chemicals and reagents
- 4.2.2 Exposure of C5a, IL-8, and pneumolysin on the 98 metals
- 4.2.3 Neutrophils
- 4.2.4 Spectrofluorimetric measurement of cytosolic Ca2+
- 4.2.5 Assay of neutrophil migration
- 4.2.6 NF-κB activation
- 4.2.7 Expression and statistical analysis of results
- 4.3 Results
- 4.3.1 Effects of prior exposure to the metals on C5a-, IL-8, 100 and pneumolysin-mediated alterations in neutrophil cytosolic Ca2+ concentrations
- 4.3.2 Effects of Pd2+on the leukotactic activity of C5a and IL
- 4.3.3 Activation of NF-κB
- 4.4 Discussion
- Chapter 5: Discussion
- Chapter 6: References
GET THE COMPLETE PROJECT
Investigation of the Pro-Oxidative and Pro-Inflammatory Interactions of Cobalt, Palladium, Platinum and Vanadium with Human Neutrophils In Vitro