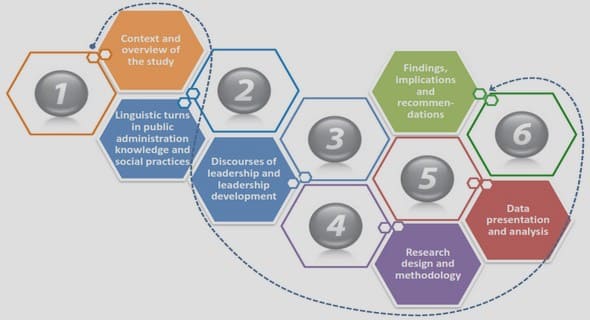
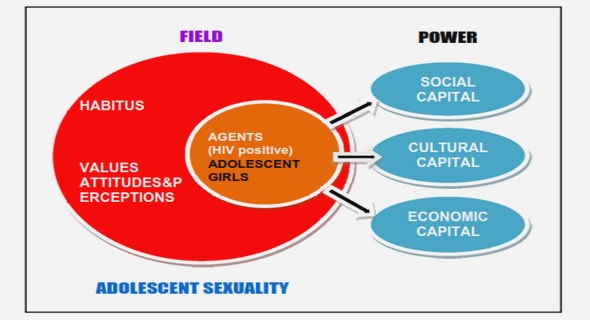
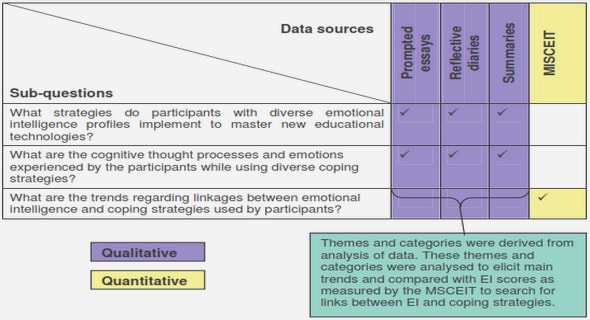
Get Complete Project Material File(s) Now! »
GENERAL INTRODUCTION
Reporter genes are commonly used in the construction of transcriptional fusions to regulatory elements of interest, and have facilitated greatly the analysis of gene expression and the study of individual promoters and their regulation. Moreover, the construction of translational fusions have also allowed for monitoring the fate of a protein of interest, e.g., degradation, subcellular localization, secretion into the extracellular milieu or translocation into other cells (Gober et al., 1995; De Kievit et al., 2001; Urbain, 2001; Møller et al., 2005; Bumann and Valdivia, 2007). The results from these analyses have found application in both basic and applied sciences. In molecular and cellular biology, reporter gene technology has contributed to improved understanding of the cellular events associated with signal transduction and gene expression, and has also provided spatial information about a particular gene product (Welsh and Kay, 1997; Tsien and Miyawaki, 1998; Veening et al ., 2004; Koga et al ., 2006).
In biotechnology, reporter gene technology has been used for the identification of novel promoters that can be used to establish improved expression systems for high-level production of recombinant proteins (Cagnon et al., 1991; Herbst et al., 1994; Serrano-Heras et al., 2005; Miksch et al., 2006). A reporter gene can be defined as a gene with a readily measurable phenotype that can be distinguished easily over a background of endogenous proteins (Wood, 1995; Naylor, 1999). Generally, such reporters are selected on the basis of the sensitivity, dynamic range, convenience and reliability of their assay (Bronstein et al., 1994; Wood, 1995; Schenborn and Groskreutz, 1999). Various reporter genes have been used for molecular genetic analyses and their encoded products all share the ability to glow, to fluoresce or to be assayed colorimetrically (Naylor, 1999; Hautefort and Hinton, 2000). However, none of the reporter systems used is universally applicable, since each reporter has its own advantages and disadvantages that may limit its use in some cells of model organisms and in certain types of studies.
Therefore, studies aimed at modifying the widely used reporter systems (Hautefort et al., 2003; Choe et al ., 2005), extending the methods used for measuring reporter activity (Thibodeau et al., 2004; Hampf and Gossen, 2006) and screening of new reporters (Post and Luebeke, 2005; Schmidt et al., 2005) are continuing. Similar to alkaline phosphatase, which has been used successfully as a reporter in both genetic analyses and in enzyme immunoassays (Reuber et al., 1991; Manoil, 2000), bacterial non-specific acid phosphatases (NSAPs) may also be exploitable as reporters in molecular biology. Bacterial non-specific acid phosphohydrolases (NSAPs) are secreted enzymes, produced as soluble proteins associated with the periplasmic space or as membrane-bound lipoproteins.
They generally dephosphorylate a broad array of substrates and have optimal catalytic activity at an acidic to neutral pH (Rossolini et al ., 1998). Many bacterial acid phosphatases function as scavengers of organic phosphoesters (Beacham, 1979), while some of them participate in an assortment of essential biological functions, including the regulation of metabolism, energy conversion and signal transduction (Stock et al., 1995; Klumpp and Krieglstein, 2002). Indeed, some NSAP-encoding genes have already been exploited as targets for insertional inactivation in cloning vectors that allow direct identification of recombinants. Using these vectors, recombinants were easily identified on the basis of their acid phosphatase-negative phenotype, while clones containing non-recombinant vector exhibited a phosphatase-positive phenotype (Burioni et al., 1995; Thaller et al., 1998a). In this review of the literature, information pertinent to reporter molecules and bacterial non- specific acid phosphatases (NSAPs) will be discussed, as they are both closely related to the aims of this study.
PROKARYOTIC GENETIC REPORTER SYSTEMS
The choice of a reporter system is determined by a number of criteria. An important consideration in this regard, is the availability of simple and sensitive methods for the quantitative and qualitative determination of reporter protein activity against the background of other cell components (Wood, 1995; Naylor, 1999). Other important properties include the ability to follow dynamic changes in gene expression (in this case, important characteristics are half-life and/or duration of protein maturation), the size of reporter gene and its product (for convenience of cloning and analysis of reporter protein expression), the absence of activities similar to that of the reporter protein in the cells under study, as well as the ability of the reporter protein to tolerate other proteins fused at its N- and C-termini without a loss of activity (Jefferson et al., 1986; Jefferson et al., 1987; Joyeux et al., 1997; Le Loir et al., 1998; Langella et al., 1999; Naylor, 1999). A variety of reporter gene systems is available and has been used extensively for analysis of different aspects of gene expression. The advantages and limitations of selected reporter genes are summarized in Table 1.1, while commonly used reporter genes are discussed in greater detail in the sections below.
TABLE OF CONTENTS :
- ACKNOWLEDGEMENTS
- DECLARATION
- SUMMARY
- LIST OF ABBREVIATIONS
- LIST OF FIGURES
- LIST OF TABLES
- RESEARCH COMMUNICATIONS
- CHAPTER ONE LITERATURE REVIEW
- 1.1 GENERAL INTRODUCTION
- 1.2 PROKARYOTIC GENETIC REPORTER SYSTEMS
- 1.2.1 Genes encoding substrate cleavage enzymes
- 1.2.2 Antibiotic resistance genes
- 1.2.3 Bioluminescence genes
- 1.2.4 Green fluorescent protein genes
- 1.2.5 Export-specific reporter genes
- 1.3 BACTERIAL NON-SPECIFIC ACID PHOSPHOHYDROLASES
- 1.3.1 Qualitative and quantitative assays for studying bacterial non-specific acid phosphatase enzyme activity
- 1.3.1.1 Plate screen assays
- 1.3.1.2 Zymogram assays
- 1.3.1.3 In vitro enzyme assays with crude preparations and purified enzymes
- 1.3.2 Classification of bacterial non-specific acid phosphatases
- 1.4 CLASS A BACTERIAL NON-SPECIFIC ACID PHOSPHATASES
- 1.4.1 PhoN acid phosphatase of Salmonella enterica serovar Typhimurium
- 1.4.2 PhoN and Apy acid phosphatases of Shigella flexneri
- 1.4.3 PiACP acid phosphatase of Pretovella intermedia
- 1.4.4 Other class A acid phosphatases
- 1.5 CLASS B BACTERIAL NON-SPECIFIC ACID PHOSPHATASES
- 1.5.1 AphA acid phosphatase of Salmonella enterica serovar Typhimurium
- 1.5.2 NapA acid phosphatase of Morganella morganii
- 1.5.3 AphA acid phosphatase of Escherichia coli
- 1.5.4 Other class B acid phosphatases
- 1.6 CLASS C BACTERIAL NON-SPECIFIC ACID PHOSPHATASES
- 1.6.1 OlpA acid phosphatase of Chryseobacterium meningosepticum
- 1.6.2 e(P4) and HppA acid phosphatases of Haemophilus influenzae
- 1.6.3 Other class C acid phosphatases
- 1.7 STRUCTURAL ANALYSIS OF BACTERIAL NON-SPECIFIC ACID PHOSPHATASES
- 1.7.1 PhoN and AphA acid phosphatases of Salmonella enterica serovar Typhimurium
- 1.7.2 AphA acid phosphatase of Escherichia coli
- 1.7.3 e(P4) acid phosphatase of Haemophilus influenzae
- 1.8 AIMS OF THIS INVESTIGATION
- 1.9 REFERENCES
- CHAPTER TWO CHARACTERIZATION O F A PHOSPHATASE S ECRETED B Y Staphylococcus aureus strain 154 AND CLASSIFICATION AS A NE W MEMBER O F T HE BACTERIAL CLASS C FAMILY OF NON-SPECIFIC PHOSPHATASES
- 2.1 INTRODUCTION
- 2.2 MATERIALS AND METHODS
- 2.2.1 Bacterial strain and characterization of the bacterium
- 2.2.2 Culture conditions and concentration of extracellular proteins
- 2.2.3 Enzyme activity assay
- 2.2.4 Influence of pH and temperature on enzyme activity
- 2.2.5 Protein determination
- 2.2.6 SDS-polyacrylamide gel electrophoresis and zymographic detection of phosphatase activity
- 2.2.7 Effects of metal ions and other reagents on phosphatase activity
- 2.2.8 Amino-terminal amino acid sequencing
- 2.2.9 Recombinant DNA methodology
- 2.2.10 DNA sequencing and sequence analysis
- 2.2.11 Sequence accession number
- 2.3 RESULTS
- 2.3.1 Staphylococcus strain identification
- 2.3.2 Characterization of the phosphatase enzyme
- 2.3.3 Properties of the phosphatase
- 2.3.4 Effect of additives on acid phosphatase activity
- 2.3.5 Sequence analysis of the gene and acid phosphatase from S. aureus
- 2.3.6 Identification of SapS homologues by sequence comparisons
- 2.4 DISCUSSION
- 2.5 ACKNOWLEDGEMENTS
- 2.6 REFERENCES
- CHAPTER THREE INACTIVATION O F T HE M AJOR CE LL WALL P ROTEASE GENE ( wprA) O F Bacillus halodurans BhFC01 AND EVALUATION OF sapS AS A R EPORTER GENE FOR HOST STRAIN EVALUATION AND CELL SURFACE DISPLAY
- 3.1 INTRODUCTION
- 3.2 MATERIALS AND METHODS
- 3.2.1 Bacterial strains, plasmids and culture conditions
- 3.2.2 Recombinant DNA techniques
- 3.2.3 Inactivation of B. halodurans BhFC01 major cell wall protease gene (wprA)
- 3.2.3.1 Construction of integrative plasmid pSEC-WprA−
- 3.2.3.2 Generation of mutant B. halodurans BhFC01 strains by in vivo recombination
- 3.2.4 Construction of surface display reporter plasmid pNW33-SC
- 3.2.5 Zymographic detection of protease activity
- 3.2.5.1 Protein sample preparation
- 3.2.5.2 SDS-PAGE and zymography
- 3.2.6 Amino-terminal amino acid sequencing
- 3.2.7 Acid phosphatase enzyme activity assays
- 3.2.7.1 Protein sample preparation and protein concentration determination
- 3.2.7.2 In vitro enzyme activity assays
- 3.2.8 Zymographic detection of acid phosphatase activity
- 3.2.8.1 Protein sample preparation
- 3.2.8.2 SDS-PAGE and zymography
- 3.3 RESULTS
- 3.3.1 Inactivation of the wprA cell wall protease gene of B. halodurans BhFC
- 3.3.2 Evaluation of heterologous protein production in B. halodurans strains BhFC and BhFC04 with sapS as a reporter gene
- 3.3.3 Evaluation of surface display in B. halodurans strains BhFC01 and BhFC with sapS as a reporter gene
- 3.4 DISCUSSION
- 3.5 REFERENCES
- CHAPTER FOUR
- 4.1 INTRODUCTION
- 4.2.1 Bacterial strains and growth conditions
- 4.2 MATERIALS AND METHODS
- 4.2.2 Recombinant DNA techniques
- 4.2.3 Plasmids
- 4.2.4 Protein sample preparation and protein concentration determination
- 4.2.5 Qualitative, quantitative and zymographic detection of phosphatase activity
- 4.2.6 Amino-terminal amino acid sequencing
- 4.3 RESULTS
- 4.3.1 Expression of the S. aureus sapS gene in E. coli, B. subtilis and B. halodurans
- 4.3.2 Evaluation of heterologous promoters and signal sequences with sapS as a reporter gene in E. coli CU
- 4.3.3 Evaluation of heterologous promoters and signal sequences with sapS as a reporter gene in B. subtilis
- 4.3.4 Evaluation of heterologous promoters and signal sequences with sapS as a reporter gene in B. halodurans BhFC
- 4.4 DISCUSSION
- 4.5 REFERENCES
- CONCLUDING REMARKs