Get Complete Project Material File(s) Now! »
The complex taxonomic history
Over the years, the taxonomy of the genus Arenaria and its allied genera has been controversial and considered unsatisfactory. For the first time, Linnaeus (Linnaeus 1737) defined this genus in the thirties of the eighteenth century. Many detailed studies have focused on the circumscription of the generic and infra-generic boundaries in Arenaria, that consistently has a confusing history. Certainly, these studies included two concepts: one held by Fernald, a wide point of view, maintaining the genus Arenaria as a whole unit. Fernald and several American botanists did not consider the different characters (valves of the capsule, foliage, ovary, inflorescence and seeds) sufficiently to demonstrate the segregated genera as distinct from Arenaria sensu stricto (Fernald 1919). Whereas the other concept divides the group Arenaria into many genera and subgenera, many botanists have adopted this strict generic concept. For example, Bentham and Hooker have recognized 11 subgenera, including the subgenus of Arenaria s. str., and proposed up to 130 species in the genus Arenaria (Bentham & Hooker 1862). Furthermore, in 7 the first taxonomic revision of Arenaria, Williams defined seven subgenera and 34 sections and subsections, which grouped 168 species (Williams 1898). Likewise, there are more than 300 species of Arenaria in the world belonging to ten subgenera and 24 sections, according to (Zhou 1995). Recently, in the last thirteen years, several authors defined a new key for the genus Arenaria and new species in Iran and in Turkey (Aytaç & Duman 2004); (Fadaie et al. 2010); (Fadaie 2013). In addition, Fior and Karis have suggested that the seed strophiole should be considered whendetermining keys to separate Moehringia L. from Arenaria (Fior & Karis 2007). However, McNeill’s viewpoint has more clearly outlined by defining the criteria of generic and infra-generic classification of Arenaria. Seven genera, and ten subgenera had been distinguished by McNeill, including the subgenus of Arenaria, that has also subdivided into several sections and series (McNeill 1962). Despite the difficulties in selecting the characters to determine the limitations, there are exceptions in a limited area, for example, in Europe (Fernald 1919), where the lines separating the Arenaria genus complex (Arenaria, Moehringia and Minuartia) are well defined by McNeill (1962).
The life forms and cycles
As we mentioned above, the geographical distributions differ extremely in genus Arenaria. Therefore, the members of this genus can be grouped into different life-forms and cycles. In general, the species of genus Arenaria are either annual or perennial herbs, sometimes biennial. However, very few detailed studies have focused on plant growth forms for this genus, McNeill has worded its life form spectrum in a way, which was categorized as plant habit. Some of McNeill’s categories are small annuals, xerophytes of Mediterranean, densely pulvinate alpines and spiny sub-fruticose (i.e., buds that are protected by dead plant remains) perennials (McNeill 1962). Raunkiaer (1934) has classified plants according to the place where the growth point is located during the less favorable seasons. The buds of Chamaephytes are on persistent shoots near the ground and the Hemicryptophytes have their dormant buds in the upper crust of the soil, just below the surface (Raunkiaer 1934). The Chamaephytes reach their highest density rates within 1400 2000m (Giménez et al. 2004), thus, we can understand that the Arenaria tomentosa Willk. (endemic species in the south of the Iberian Peninsula) is chamaephytes (Fig. 4). Furthermore, Arenaria polytrichoides Edgew is a long-lived perennial herb that forms cushion chamaephytes (Yang et al. 2010). A cushion chamaephyte is a life form in harmony with the alpine conditions.
The threat and protection
The European Red List of Vascular Plants defined the status of many species of Arenaria such as A. nevadensis, which is Critically Endangered and A. humifusa, which is Near Threatened (Bilz et al. 2011) (see Figure 6) Additionally, the status of Arenaria cinerea DC., is determined as vulnerable (VU) in Provence-Alps-Côte d’Azur (France) as mentioned in the Red Book of endangered flora in France (Olivier et al. 1995). There are also others national sources, such as the Red List of endangered species in Switzerland (ferns and flowering plants), that mentioned the status of two species of Arenaria, one of which is Near Threatened (A. bernensis) and the other is Regionally Extinct in Switzerland (OFEV 2011). Although IUCN is the best scientific tool, there are some cases with increasing knowledge of taxa, where the re-evaluation of its status is necessary, such as Arenaria provincialis Chater and Hallyday, which has reevaluated by (Véla et al. 2008). Its new status is “Near Threatened” (NT), instead of the ancient category “Rare” (R).
Intraspecific polyploidy and conservation
The study of intraspecific genome size variation can reveal inter- and even intra-populational variations in ploidy (Siljak-Yakovlev et al. 2010). The phenomenon of intraspecific ploidy variation is prevalent (Keeler 2004; Suda et al. 2007; Duchoslav et al. 2010; Realini et al. 2015), and its frequency has recently been estimated to be 12–13% for angiospermes (Wood et al. 2009). However, less attention has been paid to this phenomenon when conservation priorities are being set and it remains unaddressed in plant conservation programmes. For example, two-thirds of threatened or endangered plants in the USA are known to exhibit intraspecific chromosome number variation. yet this has been taken into account for very few species in conservation plans to our knowledge (Severns & Liston 2008; Delaney & Baack 2012). To avoid the risk of omitting information on ploidy levels in restoration measures, it would be desirable to first determine the existence of potential cytotypes. Our study identified different ploidy levels in native populations of threatened and endangered species and highlighted the importance of this form of genetic variation in plant conservation. In programmes aimed at reinforcement of populations, mixing of individuals with different ploidy levels should be stringently avoided. Here, we might consider all the populations with different ploidy levels, such as the alpine and pre-alpine populations (Mt Lure, Mt Cheval Blanc, the Mt Chaudière and Mt Raton) also, the Mt Chasseral population in the Swiss Jura, as distinct ESUs. Moreover, we could define the diploid populations presenting intraspecific variation in monoploid genome size from Pas de l’Ai in the Mt Sainte Baume chain in France and the Mt Sagra in Andalusia as distinct ESUs.
Table of contents :
1. Chapter I: General introduction
1.1 Polyploidy in plants – a brief history
1.2 Evolutionarily significant units (ESUs)– a confuse history
1.3 Arenaria L. – a complex history
1.4.1 The complex taxonomic history
1.4.2 The distribution and ecology
1.4.3 The life forms and cycles
1.4.4 The morphological description
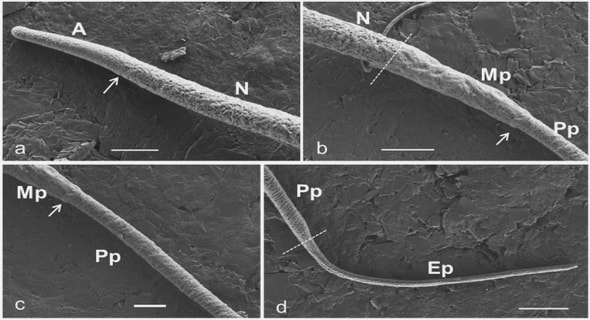
1.4.5 The seeds
1.4.6 The threat and protection
1.4.7 The phylogeny
1.4 Conclusion of the introduction
2. Chapter II: Genetic variation within the species complex of Arenaria grandiflora L.
2.1 Introduction to Chapter II
2.2 Paper 1: Intraspecific DNA content variability: consequences for conservation
3. Chapter III: Germination patterns within Arenaria grandiflora L. species complex
3.1 Introduction to Chapter III
3.2 Paper 2: Seed germination of Arenaria grandiflora L. species complex (Caryophyllaceae)
4. Chapter IV: Morphometric variation within the species complex of Arenaria grandiflora L.
4.1 Introduction to Chapter IV
4.2 Paper 3: Leaf shape discriminates among populations with different ploidy levels and from
different environments in the Arenaria grandiflora (Caryophyllaceae) species complex
5. Chapter V: General Conclusion