Get Complete Project Material File(s) Now! »
Bad tenants: Female sheet-web spiders (Cambridgea foliata) lose feeding opportunities when cohabiting with males
In review at Journal of Arachnology
ABSTRACT
In web building spiders, females are often too widely distributed across the landscape for males to monopolise more than one mate. Consequently, males seek females one at a time and may cohabit with females in their webs. Pre-copulatory cohabitation is most common in aranaeomorphs which suggests that the first male to mate with a female will have a greater share of paternity than any subsequent mates (first sperm precedence). However, pairs of adult New Zealand sheet-web spiders (Cambridgea foliata) cohabit for longer than required to achieve copulation. This is counter-intuitive as it suggests that males defend females which they have already copulated with in lieu of seeking additional mating opportunities. To investigate the costs and benefits of extended cohabitation on male and female Cambridgea we conducted surveys of webs of solitary and paired males and females. We found that solitary spiders of both sexes consistently position themselves in the centre of their webs but that when in pairs, females are displaced from the webs by males and will frequently leave the web altogether. Males in pairs would respond to vibrations simulating prey, while females would not respond. This strongly suggests that extended cohabitation should be costly for females. By contrast, for males, cohabitation is a valuable foraging strategy which, combined with the advantages of mate guarding, may compensate for any lost mating opportunities due to their foregoing searching for further mates.
INTRODUCTION
Males regularly compete to ensure mating success and to increase the likelihood that their sperm will be used by females to fertilise their eggs (Parker 1970). This competition can manifest in a variety of ways including defence of mates or of resources, and scramble competition. In the former, males compete through agonistic contests or displays to defend spatially clustered females or resources important to females (Emlen & Oring 1977; Thornhill & Alcock 1983). In the latter, if females are widely distributed or are only available for a limited time, defence is not economical so males scramble to mate with as many females as possible. This seems to be a relatively common mating system within spiders (e.g. Desert spiders, Stegodyphys lineatus, (Berger-Tal & Lubin 2011); Redback spiders, Latrodectus hasselti (Kasumovic & Andrade 2009); Bowl and doily spiders, Frontinella pyramitela (Austad 1984); Golden orb spiders, Nephila plumipes (Kasumovic et al. 2007)).
However, while it is advantageous for male spiders to mate multiply over their life time, brief periods of cohabitation, a form of mate guarding, are not uncommon. In spiders, mate guarding males will share nesting chambers or webs, or simply remain near females’ webs. Male spiders predominantly cohabit with subadult females with records for 161 species in which adult male aranaeomorphs cohabit with conspecific juvenile females (Jackson 1986). In these cases, males often mature earlier than females, find a female one or two moults from maturity and remain with her until she matures.
This predominance of males cohabiting with subadult females is logical provided that the first male to mate with a female receives some advantage over subsequent males. If that male alone mates with the female and she oviposits shortly after or becomes unreceptive, then he will secure 100% paternity for her clutch. However, while cohabitation with subadults is more common, post-copulatory cohabitation with adults does occur even at the cost of additional mating opportunities (Parker 1974). Alcock (1994) identifies several alternative hypotheses to explain the evolution of prolonged male-female associations following copulation. Mate guarding may prevent females from accepting additional copulations (Thornhill 1948b) and favouring the ejaculates of subsequent males (Eberhard 1991). In species that exhibit “last male sperm precedence” in which subsequent mates receive a greater share of paternity, it is valuable for males to guard their mates (Austad 1984). In some cases guarding occurs until oviposition (e.g. cellar spiders, Pholcus phalangiaoides, Schaefer & Uhl 2003) or until females enter a refractory period during which they are unreceptive to further mating (e.g. marbled cellar spiders, Holocnemus pluchei, Calbacho-Rosa, Córdoba-Aguilar & Peretti 2010).
Males may also associate for longer in order to mate multiply and increase their share of paternity (Simmons 2001). For example, male leaf-curling spiders (Phonognatha graeffei) improve fertilisation success through multiple, prolonged copulations and for this reason cohabit with adult females for several days (Fahey and Elgar 1997). While only a short period of pre-copulatory cohabitation may be needed to ensure a single copulation, longer periods of cohabitation may maximise fertilisation success for the male.
Nevertheless, cohabitation should be costly to males who forgo seeking additional mates (Birkhead & Møller 1992; Fryer, Cannings & Vickers 1999; Harts & Kokko 2013). However, these “costs” assume not only that there are a large number of receptive females available elsewhere but also that, if they should locate additional mates, males would not meet any resistance from other guarding males (Harts & Kokko 2013). Furthermore, remaining in a female’s web may provide additional benefits not directly related to reproduction such as safety. Austad (1984) remarks that post-copulatory mate guarding should only be advantageous if moving between webs, and finding a newly moulted female, is risky. This risk of travelling has been demonstrated in several spider species in which less than a quarter of males observed in a female’s web are found again in the web of another female (e.g. golden orb web spider, Nephila clavipes, Christenson & Goist 1979; bowl and doily spider, Frontinella pyramitela, Austad 1984; redback spider Latrodectus hasselti, Andrade 2003). Predation by other invertebrates is likely to be greatest source of mortality (Andrade 2003). Consequently, it can be advantageous for males to maximise their reproductive output with a resident female rather than risk searching for another.
Cohabitation may also provide males with greater feeding opportunities. Suter and Walberer (1989) found that male bowl and doily spiders (Frontinella pyramitela) feed on prey caught in females’ webs and suggest that this could be why cohabitation can go on for several days, far longer than is necessary for only courtship and copulation. They suggest that this may compel the female to eventually expel her tenant. Furthermore, Erez, Schneider & Lubin (2005) demonstrated clear benefits of cohabitation for males at the expense of resident female desert spiders, Stegodyphus lineatus. When they provided food to webs containing cohabiting pairs, the males would gain in condition while females would not. In species that depend on capture webs to feed, foraging in a resident female’s web is an ideal way for males to prepare for searching for his next mate (Austad 1983). Apart from these studies, there are no other examples that we are aware of which test whether males gain feeding benefits while cohabiting on female webs.
Cambridgea foliata (Desidae) are arboreal spiders distributed throughout the North Island of New Zealand. They are nocturnal and build three-dimensional sheet-webs in native forest which males and females can be seen sharing. The webs consist of a non-sticky, thick horizontal mainsheet, which the spiders run along the underside of, anchored from below and with many knock-down threads above the mainsheet. The rear of the web tapers into a silken tunnel or “retreat” which may extend under the bark of trees or into vacated burrows which the spider resides in during the day. During the summer, males mature in their natal webs before leaving to wander in search of female webs. Once a male finds a female’s web he may cohabit with her. McCambridge (2017) found that C. foliata will cohabit for between one day to about a fortnight with a small number of males cohabiting with subadult females for more than 20 days. Based on the author’s data and our own observations, we expect that one to two days is the most common duration of cohabitation. During this time, the cohabiting male will defend the female’s web by engaging in ritualised fights with intruding males (pers. obs). When males fight in a female’s web, we have often observed the resident female departing the mainsheet and sitting either on one of the anchoring threads or off the web entirely. This suggests that one male’s presence in females’ webs, or a succession of cohabiting males, may entail some foraging cost to females. However, we have not observed any instances of sexual cannibalism or fighting between males and females. Adults of both sexes are similar in size.
Courtship and mating behaviours have not been observed in this species. However, as most spiders take only a few minutes to copulate (Fahey & Elgar 1997), it is reasonable, barring the use of mating plugs or genital mutilation (reviewed in Huber 2005), to expect that cohabiting pairs have the opportunity to mate more than once. Males often cohabit with juvenile females which is consistent with Austad’s (1984) predictions for case of first sperm precedence but have also been observed cohabiting with adults. As C. foliata are entelegyne spiders with “conduit” or “one-way” type copulatory ducts, we may expect them to exhibit first sperm precedence. However invertebrate copulatory and fertilisation duct morphology is highly variable which can generate significant variation in the strength of first or last sperm precedence (Uhl 2000). For this reason, we do not assume whether this species exhibits first or last sperm precedence.
Given that male C. foliata cohabit with females for longer than required for a single copulation, in the present study we aim to assess the impact of males cohabiting with females by comparing positions of male and female spiders on their webs when alone and when cohabiting, and (2) comparing responses of males and females to simulated prey. If we assume that the probability of prey interception is approximately equal at all points on the mainsheet then the centre of the mainsheet (which we call the “hub”) should be the optimal location for the spider to sit as it is approximately equidistant to all edges of the web and to the retreat where these spiders hide if disturbed. For this reason, we predict males and females will most commonly position themselves in the hub when alone. When in pairs, we expect that both sexes will spend less time in the hub compared to when alone but that males will spend more time in the hub than females as the mainsheet serves as the arena on which males fight each other. Due to males and females sharing the web when in pairs, we expect that both will be less likely to approach a simulated prey item compared to when alone. Another objective is to describe courtship interactions on females’ webs, which have not been described before and may be useful for understanding mate guarding in this species.
METHODS
Courtship observations
In addition to making observations of male-female interactions in the field, we collected juvenile C. foliata for a lab population which included females with a known reproductive history. Each spider was housed in a 30×30×60.1 cm mesh enclosure. A wooden retreat was attached to the upper rear corner of the enclosure. We modelled these on wētā enclosures devised by Bowie et al. (2006). They consist of a (45 × 45 × 150 mm) block of untreated wood with a furrow (18 ×18 mm) cut two-thirds the length of the block which was placed against the mesh wall. A black polyethylene flap was attached to the outside of the cage, covering the retreat. We fed the spiders every two days on a mixture of meal worms (Tenebrio moliter), blow flies (Lucilia sericata) and locusts (Locusta migratoria). Spiders were misted with water three times a day.
By the time females reached maturation their webs were dense enough to support multiple spiders. In order to observe courtship interactions, we introduced a random male to a random female’s web and recorded subsequent behaviours for up to 3 hours.
Cohabitation surveying
We surveyed C. foliata webs at night in Matuku Forest and Bird reserve in West Auckland (36° 51′ 48.3″S 174° 28′ 47.7″E). We observed webs on either side of the walking track across the 2015/2016 and 2016/2017 summer seasons. We could ensure that we did not observe the same individuals across the separate seasons as in 2015/2016 the spiders were subsequently collected for other research. Meanwhile in the 2016/2017 season we ensured that we did not repeat observations of the same spiders by not visiting the same trees.
When we found an adult spider in a web we made a note of sex, location in the web and whether the spider was solitary or in a pair. The “locations” we included were “retreat” meaning at the opening retreat or on the mainsheet immediately outside the retreat, the “hub” referring to approximately the centre of the mainsheet, the “web” referring to all remaining parts of the mainsheet and “off” indicating that the spider was either on the knock-down threads, on the guying threads or just off the mainsheet on the substrate within 10cm of the web.
To examine whether spiders changed how they responded to a prey stimulus when in a pair compared to when alone, we provided an artificial stimulus simulating prey movement in the web. In order to standardise the stimuli we used a Wittner tuning fork with a pitch A440 (440 Hz). We had previously observed that C. foliata females consistently approach a vibrating tuning fork when touched to the mainsheet of the web and will attempt to bite it before realising that it is not a prey item. In the current study, we would strike the tuning fork and touch the tip lightly approximately 20cm from spider’s location. This controlled for differences in web size which varied considerably (mean area = 3169.48 cm2, SD = 1220.63).
We recorded whether the spider(s) approached, moved away from or did not respond to the stimulus. As most individuals either responded within 5 seconds of the stimulus being applied or did not respond for more than 30s or with additional applications of the stimulus, we did not record latency.
We surveyed the webs of 62 solitary females, 34 solitary males and 47 adult pairs. We collected response data for 35 solitary females, 22 solitary males and 23 pairs. When analysing spider locations and responses, we excluded 11/62 of the webs with solitary females, 3/34 webs with solitary males and 5/47 webs with pairs in cases when spiders were feeding. We kept this data separate to our other location data as a prey item falling into the web would draw one or both spiders away from their original location and therefore would not be representative of the “preferred” location.
Statistics
We conducted a χ2 test for homogeneity of variance on contingency tables for female locations depending on whether they were alone or in a pair. When tabulating female responses to the stimulus, our expected count values for our contingency tables violated the assumptions for a χ2 test of homogeneity. As the vast majority of responses were either that the female would approach or not respond and very few would move away from the stimulus, we pooled negative responses and no response together. The resulting contingency table for whether females approached the stimulus (with binary responses of yes or no) did not violate the assumptions forχ2
We calculated odds ratios for the probability that females would take up certain positions when in pairs compared to when alone, and for the probability that they would approach a stimulus as opposed to move away or not respond. We conducted these same tests for males. All analyses were conducted in R version 3.2.0. (R core team 2015).
RESULTS
Courtship
We observed 10 instances of courtship. Within these observations, 6 males entered a copulatory posture. When males first enter webs occupied by females they may shake the web using their whole body. The female orients towards him and, soon after, approaches him. She touches him with her forelegs and then may retreat a short distance in the direction of the retreat. The male may continue to interact with the female or move past her and enter the retreat. The interactions we observed lasted for 93.8s (SD = 168.3s). In one case, a male courted a female for 615s.
Often at the entrance of the retreat, the male shakes the web and dorso-ventrally flexes his abdomen. If the female approaches he will drum on the web with his first and second pairs of legs. The female may angle her cephalothorax away from the web allowing the male to approach and place one palp over her epigyne (Fig. 1). When the male moved out of this posture, he would either remain on the web a short distance from the female or return to the retreat.
Contents
ABSTRACT
DEDICATION
ACKNOWLEDGEMENTS
1. GENERAL INTRODUCTION
1.1. Sexual selection and intrasexual competition
1.2. Scaling relationships in animal weaponry
1.3. The role of weaponry in animal contests
1.4. The evolution of exaggerated weaponry and positive
1.5. Spiders as a study system
1.6. Outline of thesis
2. THE ROLE OF EXAGGERATED MALE CHELICERAE IN MALE-MALE CONTESTS IN SHEET-WEB SPIDERS
2.1. Abstract
2.2. Introduction
2.3. Methods
2.4. Results
2.5. Discussion
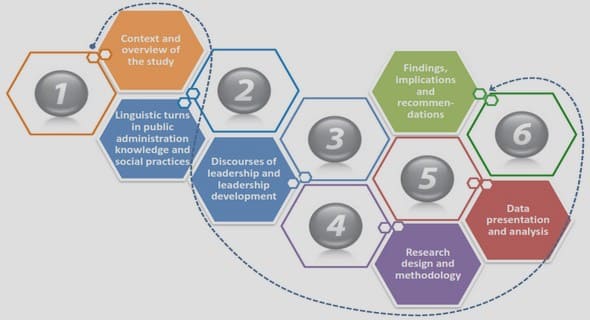
3. BAD TENNANTS; FEMALE SHEET-WEB SPIDERS LOSE FEEDING OPPORTUNITIES WHEN COHABITNG WITH MALES
3.1. Abstract
3.2. Introduction
3.3. Methods
3.4. Results
3.5. Discussion
4. THE EVOLUTION OF POSITIVE ALLOMETRIES IN NEW ZEALAND SHEET-WEB SPIDERS
4.1. Abstract
4.2. Introduction
4.3. Methods
4.4. Results
4.5. Discussion
5. HARDER, BETTER, FASTER, STRONGER; WHAT IS THE ADVANTAGE OF EXAGGERATED WEAPONRY?
5.1. Abstract
5.2. Introduction
5.3. Methods
5.4. Results
5.5. Discussion
6. GENERAL DISCUSSION
6.1. Key findings
6.2. Synthesis
6.3. Implications
REFERENCES
GET THE COMPLETE PROJECT
Exaggerated male weaponry in New Zealand sheet-web spiders (Cambridgea)