Get Complete Project Material File(s) Now! »
Poplar hybrids
Spontaneous hybrids have been used in commercial culture for several decades. In addition to that, geneticists bred hybrids that were deployed in poplar cultivation throughout the world. The most common of these commercial hybrids belong to Populus # euramericana, which is the cross Populus deltoides (#) ! Populus nigra ($). To date, tree breeders continue producing new hybrid cultivars using microculture and biotechnology techniques to cross mating barriers.
In France, apart from a few genotypes of Populus deltoides, Populus trichocarpa and Populus alba, poplar cultivation is dominated by (i) euramericana hybrids (Populus deltoides # nigra (ii) interamericana hybrids, stemming from the cross Populus deltoides # trichocarpa. Interamericana hybrids have been present in the French national catalogue since 1982. Over the past 20 years, the resistance of these genotypes to fungal attacks drove the relative share of these two types of hybrids in french plantations. Interamericana hybrids showed remarkable hybrid vigour. Thus, between 1987 and 1997, interamericana hybrids, represented mainly by ‘Beaupré’ left very little room for euramericana cultivars. Nevertheless, these cultivars grown in dense plantations suffered severe rust attacks (Melampsora larici-populina Kleb.) after 1997 due to their low quantitative resistance (Dowkiw & Bastien 2007). They were less and less used thereafter. Until 1998, sales of interamericana hybrids increased meagerly but thereafter decreased sharply in favour of the euramericana hybrids (Breton 2000; Paillassa 2001; Balzinger & Ginisty 2002). For almost 20 years, a strong increase in the relative proportion of Euramericana cultivars growing in France, especially for the clone ‘I214’ since 2000 was detected by FCBA (Forêt, Cellulose, Bois-construction, Ameublement; Thivolle-Cazat 2002). This shows that after a fall in interest for Euramericana cultivars between 1987 and 1997, farmers are showing a renewed interest in these hybrid poplars (see Figure 4 for its distribution in 2003).
Poplar cultivation
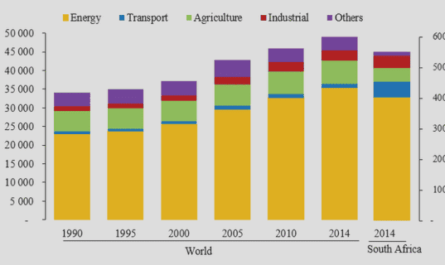
Traditionally, poplar has been cultivated in France since the 18th century. Due to import ban against trembling aspen for matches and okoume for plywood, poplar cultivation has regained farmer’s interest. Poplar cultivation was once again boosted after the creation of « The French Poplar Commission » in 1942, which became the « International Poplar Commission IPC » in 1947. It was a substantial initiative of France to promote poplar cultivation in collaboration with FAO (Food and Agriculture Organization of the United Nations). Since then, additional 28 countries have joined in and have enlarged the list of 8 pioneer members who responded to the call of attending the poplar week in April 1947. The tasks of IPC are multiple: to study the scientific, technical, socio-economical aspects of poplar culture; to promote the exchange of ideas and materials among researchers, producers and users; to create joint research programs and stimulate organization of meetings (Viart & Fugalli 1997; Viart 1999). In France, due to the long efforts of IPC and other forestry related organizations, the area under poplar cultivation increased from 100000 ha in 1939 to almost 185100 ha in 2003 (Table 3), which increased to 235000 ha in 2008.
Region wise, Picardie (32000 ha), Champagne-Ardenne (27000 ha), Aquitaine (25000 ha) and Centre along with Pays de la Loire (44000 ha) share most of the area under poplar cultivation (Table 3 & Figure 4). Such a large area makes poplar the third highest wood producing species in France after oaks and beech in 2003 and the second one in 2008 after oaks (1.5 million m3 of wood per year, FAO 2008).
Poplar wood is very versatile and widely used. It is light in weight (specific gravity 0.3 to 0.4) because of its porous anatomy. The wood is soft, creamy white in colour (except for a dark, pale brown heartwood or wood core), straight-grained and uniform in texture. Some species readily produce tension wood characterized by a low cell wall lignification which results in a woolly aspect of the wood. Wood of many poplar species displays a low strength, stiffness, shock resistance, decay resistance and bending properties. However, the wood can be worked easily with hand or machine tools. In addition, poplar wood can be easily glued, screwed and nailed. Staining can be patchy but paints and varnish are easily applied. Poplar wood is widely used to manufacture pulp and paper, peeled and sliced veneer, composition boards (including oriented-strand board), sawn lumber, crates, boxes, matches, chopsticks, poles, furniture, cabin logs, and excelsior. In 2008 in France, 32% of the harvested poplar wood volume was used for light peeling, 7.9% for veneer plywood, 24.4% for export veneer, 24.7% for sewing palettes and 11% for sawing crates and bedding.
Boxplots for the values of the discrimination against 13C (13C) recorded in the bulk leaf matter of a range of Populus # euramericana genotypes grown in a factorial experiment in the field at Orléans, during 2003 and 2004. During 2004, a moderate drought treatment was applied to half the individuals and induced a decline of13C. Values recorded during 2003 were influenced by the heat and drought stress that happened during that year. Based on Spearman’s rank correlation (rs), the genotypic ranking remained same as found in 2003. Modified from Monclus et al. 2006.
Research questions and thesis objectives
Poplars occupy an important place in the timber industry. To meet the currently increasing industrial demand, growers are bringing new areas under poplar cultivation such as in the Pays de la Loire and Garonne regions. As a result of this extension in addition to predicted climate change, an enhanced risk of drought events might have a negative influence on the productivity of poplar genotypes. In this scenario, the question of the effect of water supply on productivity is becoming more and more important. In this context, the concept of transpiration efficiency (total biomass accumulated with respect to water transpired) gains interest. More transpiration efficient poplar genotypes, selected on the basis of reduced water consumption (or stomatal conductance) could ensure efficient water use and a sustained production by enhancing conservation of soil water and allowing plants to remain physiologically active.
In this respect, the work done during this thesis fits the long chain of studies done on poplar genotypes during a collaboration between two laboratories, Forest Ecology and Ecophysiology (EEF, UMR 1137 INRA/Université de Lorraine) and Laboratoire de Biologie des Ligneux et des Grandes Cultures (LBLGC, UPRES-EA 1207, University of Orléans). The aim of these two labs is to identify the ecophysiological and molecular bases controlling the genetic and phenotypic variation of complex traits in trees. Traits of interest include water use efficiency (WUE) and drought tolerance. In this regard, their main focus is on the commercialized poplar cultivars like Populus # euramericana and wild type Populus nigra. The aim is to detect genetic material that is both water-use efficient and adapted to the different cultivation regions. In this context, the detection of genotypes, noticeably of Populus # euramericana, were the centre of interest (thesis dissertation of N. Marron 2000-2003, R. Monclus 2002-2006 and L. Bonhomme 2005-2009).
Poplar hybrids are known for their potential to produce high volumes of wood in short periods of time (Marron et al. 2005; Marron & Ceulemans 2006; Marron et al. 2007) but are generally sensitive to drought. Their productivity is very closely linked to water availability that can limit growth, wood quality and eventually favour the installation of pathogens (Marçais et al. 1993; Barigah et al. 1994; Pinon & Valadon 1997; Loustau et al. 2005). Therefore, it becomes a very important issue to detect genotypes that combine productivity, high wood quality and tolerance to biotic and abiotic stresses. In Populus # euramericana genotypes, previous studies have highlighted a wide range of genotypic variation for growth potential, transpiration efficiency at leaf level (estimated through 13C discrimination; see Figure 5 b; discrimination against 13C ( and productivity traits like above ground biomass (ABiom), stem diameter (dDiam) and maximum leaf area (TLAmax) as they are present on the opposite axis on the PCA (b) and large genotypic variability for productivity traits and between tested 29 genotypes (Populus deltoides # nigra). varied between 1‰ for Pannonia (Pn) and 21.51‰ for Eco-2 (E28). Reprinted from Monclus et al. 2005.
Marron et al. 2005; Dillen et al. 2008) and tolerance to moderate drought (Monclus et al. 2005; Monclus et al. 2006; Bonhomme et al. 2008). A large genotypic variability has also been found in the amplitude of the drop of productivity in response to water shortage among poplar cultivars (Gebre et al. 1994, 1998; Liu & Dickmann 1996; Chen et al. 1997; Harvey & van den Driessche 1997; Robison & Raffa 1998; Tschaplinski et al. 1998; Marron et al. 2003; Monclus et al. 2006).
These initial studies were based on the measurement of instant transpiration efficiency (TE) through leaf gas exchange measurements and of time-integrated TE using the natural abundance of stable carbon isotopes (13C) in leaf tissues (Marron et al. 2005; Monclus et al. 2005, 2006; Bonhomme et al. 2008; Dillen et al. 2008, 2009). Along with the variability detected among genotypes in the discrimination against 13C from the atmosphere to the leaf matter (13C), morphological indicators for productivity were identified such as the maximum area of single leaves and total leaf area. Furthermore, no relation was evidenced between productivity and13C (Fig. 6 a, b) both in open field experiments and under controlled environment. Under moderate drought,13C decreased but the genotypic ranking remained intact, evidenced through Spearman’s rank correlations (see Figure 5; Monclus et al. 2006). Along with this large genotypic variability for productivity and the stability of genotype ranking for13C, stomatal conductance was found to be responsible for genetic variation in13C (Monclus et al. 2006). More recently, Dillen et al. (2008) also evidenced the lack of direct relationship between stomatal traits and plant growth or13C in Populus # euramericana genotypes. Such results in various poplar genotypes (i.e., the absence or weak relationship between13C and growth) open the possibility of selecting genotypes combining high productivity and high transpiration efficiency.
In these studies, the genetic variability of TE was exclusively measured through13C (as an estimator of TE). Furthermore, experiments included very young individuals (up to 2 years) grown in a greenhouse or in open-field experiments. Extrapolating the conclusions obtained at leaf scale and concluding about stability of genotypic variability for whole plant TE can be tricky. Factors likec andw (carbon lost during respiration and water lost during night respectively) may vary between genotypes, thus the observed genotype variability in13C may not perfectly match the genotype variability in whole plant TE. Previous studies have found that variability in13C does not necessarily reflect the variability in whole plant TE (Sun et al. 1996; Picon et al. 1996a; Turner et al. 2007; Matzner et al. 2001). Thus, to select genotypes for lower 13C, we need to be sure that observed genotype differences in13C.
Table of contents :
1. INTRODUCTION
1.1. Genus Populus
1.2. Poplar hybrids
1.3. Poplar cultivation
1.4. Problematic and thesis objectives
2. STATE OF THE ART
2.1. Water use efficiency at different integration scales
2.1.1 At leaf level
2.1.2 At whole plant level
2.2. 13C discrimination as an estimator of intrinsic transpiration efficiency
2.3. Other sources of variation in 13C signals: isotopic signature of different organic compounds
2.4. Use of oxygen isotopes as estimator of stomatal conductance
2.5. 18O enrichment in the leaf
2.5.1. 18O enrichment of leaf water at the sites of evaporation
2.5.2. 18O enrichment of leaf organic matter
2.6. Dual isotopic approach (13C and 18O)
2.7. Age effect and potential causes
2.8. Sampling strategies to study age effect
2.9. Common problems in tree ring isotopic study
2.9.1. Choice of a wood component for isotopic analysis
2.9.2. Correcting chronology for trends in atmospheric 13C Transition
CHAPTER 1: Genotype differences in 13C discrimination between atmosphere and leaf matter match differences in transpiration efficiency at leaf and whole-plant levels in hybrid Populus deltoides ! nigra Transition
CHAPTER 2: Vapour pressure deficit during growth does not affect the ranking of Populus nigra genotypes for transpiration efficiency at leaf as well as at whole tree level Transition
CHAPTER 3: Time course of !13C in poplar wood: genotype ranking remains stable over the life cycle in plantations despite differences between cellulose and bulk-wood
Transition
CHAPTER 4: Age related trend in !13C does affect the genotypic ranking in 3 Populus euramericana genotypes: A synchronic approach.
GENERAL DISCUSSION AND CONCLUSION
PRESPECTIVES
REFERENCES