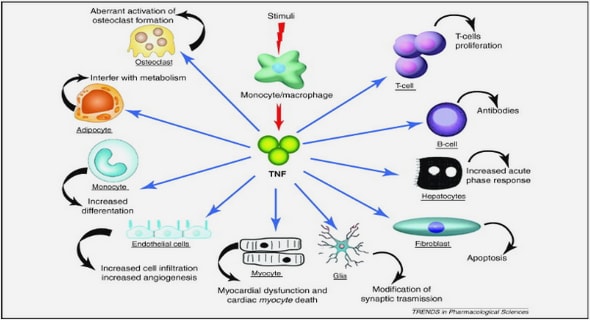
Get Complete Project Material File(s) Now! »
Chapter 3 In vitro identification of inhibitors against PfPdx1 and their potential MOA
Introduction
On the inner workings of the PLP synthase complex
The Pdx1 protein combines R5P and G3P, including ammonia provided through the hydrolysis of L-glutamine by Pdx2, to form PLP. Merging of these small molecules occurs in the Pdx1 component of the PLP synthase complex, and how this is coordinated is not entirely understood yet. The Pdx1 reaction mechanism leading to the formation of PLP is fascinating and intricately complex. Some significant findings regarding the inner working of this enzyme are briefly introduced, which alludes to peculiar functions that make it an attractive drug target.
The Pdx1 enzyme perform a multitude of different complex chemical functions including triose-and pentose-phosphosugar isomerizations, condensation of two phosphosugars, imine formation, and closing of the pyridine ring after incorporation of ammonia [119, 162]. The PfPdx1 protein is highly conserved in organisms utilising the DXP-independent B6 pathway, including P. falciparum parasites [117, 188]. From this high degree of structural conservation it can be deduced that the multiple Pdx1 active sites have an uncompromising nature, and point mutations are not readily tolerated in this gene. This could hold merit in terms of inhibitor design, since fewer incidences of rescue of resistant phenotypes due to point mutations in PfPdx1 could be expected.
Pdx1 proteins have the ability to utilise alternative substrates such as dihydroxyacetone phosphate (DHAP) and Ru5P as alternative sugars [194]. In particular PfPdx1 also has this ability [104]. DHAP and G3P are interchangeable triose sugar substrates, and it was shown that Pdx1 contain triosephosphate isomerase (TIM) activity, able to catalyze the conversion of G3P into DHAP [194]. Both R5P and Ru5P are accepted substrates in Pdx1 proteins, however, B. subtilis Pdx1 (BsPdx1) was demonstrated to have a lower Km value for R5P, which suggests this to be the preferred substrate [195]. Both R5P and Ru5P are incorporated into the final PLP molecule, and the enzyme has been proposed to have R5P isomerase activity.
As mentioned previously, the R5P binding site or P1 site is known from crystallographic evidence [157, 164]. The P2 site or PLP-binding site is located on the surface, and was shown to be solvent accessible with four water molecules complexed with PLP [163]. However, where the R5P and G3P substrates combine, and whether this occurs in the P2 site is not known [163]. A third site or P3 site in which G3P was bound, has been identified, and the G3P molecule had H-bond contacts to E116 and R164 [163]. Equivalent residues of PfPdx1 that could be involved in G3P binding are R164 and D119.
It is widely accepted that the R5P / Ru5P substrates form imines with an active site lysine as one of the first steps of the reaction mechanism [157, 161, 162]. Hanes et al. identified initial reaction 13C-nuclear magnetic resonance (NMR) peaks characteristic of an imine bond between the N atom of lysine and the C1 carbon of the R5P substrate (2 in Figure 3.1) [160]. Additional evidence to support this was shown in the PbPdx1 structure shown with bound R5P [164]. Earlier structural evidence of Ru5P bound in the TmPdx1 crystal structure has proven that the molecule is capable of forming an imine with the active site lysine, however substrate isomerisation and reshuffling of Ru5P could lead to reversion into the bound R5P state [157, 160, 194].
only the pentose substrate R5P. Formation of the I320 chromophoric species was also found to be dependent on the R5P concentration in the presence of the nitrogen source, including ammonium sulphate [162]. Systematically, incubation of R5P with Pdx1 results first in the formation of the imine- or amine-intermediate, then into I320 once the ammonium sulphate source was introduced (4 in Figure 3.1) [161, 162]. Hanes et al. proved that the I320 chromophore was a ketone intermediate species of R5P which further undergoes imine formation with G3P leading to eventual formation of PLP (5 in Figure 3.1) [160, 196]. The I320 intermediate can therefore be considered as a marker for R5P binding and an indication that R5P is effectively converted into an intermediate receptive for conjugation of G3P.
Some reshuffling of both the R5P substrate and lysine migration in the R5P-active site has been proposed; Hanes et al. showed that there is an imine migration (also called lysine migration) from the C1 of R5P (2 in Figure 3.1) to the C5 (3 in Figure 3.1) after incorporation of ammonia (there are multiple step involved) [160]. This implies that there is a state during the reaction mechanism when an early intermediate form of R5P dissociates from the internal lysine, in order to form the new C5 imine bond with the same lysine. This step could have implications for inhibitor design, and compounds that interfere with imine formation initially as well as during this step could have potential to affect the enzyme activity. A second proposal by Zein et al. suggested the possibility of an imine shuffling in TmPdx1 involving both K81 and K149 residues in TmPdx1 [157]. The R5P was suggested to be shuffled or transferred to a new site in which the K149 residue forms the imine bond (in this case with the C2 carbon). Mutagenesis of K149 did disrupt PLP formation, consistent with the notion that this residue has direct catalytic involvement [157]. The K149 in TmPdx1 was suggested to be capable of rotating, coming in close proximity with the Ru5P-K82 imine adduct Both these proposal suggest that imine formation in the R5P-active site is essential for catalytic conversion of R5P, and compounds that interfere with these steps could be valuable during drug design. Many questions remain regarding the reaction mechanism of Pdx1. The site of G3P-binding in Pdx1 has only recently been proposed and how this substrate is brought into proximity to R5P is not yet known [163]. The active site residues such as D26, K83 as well as K151 are strictly conserved in PfPdx1 suggesting a similar reaction mechanism.
Recent mutagenesis studies on the PfPdx1 protein have also highlighted important residues in dodecamer assembly as well as enzyme activity. Two triple mutants of PfPdx1 were created the DKK (D26A, K83A and K151A) mutant involved mutation of the active site residues, and the RHE (R85A, H88A and E91A) mutant involved residues involved in Pdx1:Pdx1 monomer contacts [133]. Both of these PfPdx1 mutants were not capable of maintaining dodecameric quaternary structure of the PLP synthase and existed as inactive Pdx1 hexamer and monomer structures, respectively. [133]. In the DKK mutant the residues that destabilise PfPdx1 assembly were shown to exclusively the K151A mutation [133]. The monomeric RHE PfPdx1 mutant, which also did not support Pdx2 activity, suggested that these highly conserved RHE Pdx1:Pdx1 interface residues coordinates Pdx1 assembly to form at least a hexameric Pdx1 crown [133]. The ability of these mutants to form the I320 intermediate – a marker of R5P binding – was not tested. The previously created PfPdx1 DKK and RHE mutants were therefore tested in order to establish the involvement of these residues in the catalytic process of forming PLP.
The C-terminal tail of the PfPdx1 also contributes to oligomeric assembly and substrate binding. A C-terminally truncated PfPdx1 (PfPdx1 273-301 and PfPdx1 270-301) was monomeric and this 30 amino acid unstructured region is suspected to be involved in initial substrate binding and cooperative assembly of the PLP synthase [182]. This mutant was also unable to form I320, and through long-range interaction is believed to aid in organisation of the R5P active site. Here the PfPdx1 270-301 mutant was recreated and the catalytic capabilities in terms of PLP and I320 formation were confirmed in this study. This also served as a control during analysis of other PfPdx1 mutants and their ability to form I320. Additionally, the residue located in the P3 G3P binding site was mutated to confirm this residues involvement in PLP formation as well as I320 formation. This residue was located more than 20 Å from the R5P binding site, and was not expected to affect I320 formation [163]. In the previous chapter, the identification and selection of potential inhibitors of PfPdx1 were reported. Here the effect of these compounds was tested on purified PfPdx1 in vitro. Some active inhibitors were identified and the potential mode-of-actions of these are discussed further.
Methods
Expression and purification of PfPdx1
Plasmids of wild-type P. falciparum Pdx1 and Pdx2, as well as K83A, K151A, RHE, ERR, DKK PfPdx1 mutants, cloned into the pASK-IBA3 vector, were obtained from Knöckel et al. [132]. Plasmids were transformed into the E. coli BLR (DE3) expression cell line using heat shock transformation. PfPdx1 plasmid DNA (1 µL or 10 ng) was added to 100 µL heat shock competent cells (prepared as described below), allowing incubation on ice for 30 min. Cells were subjected to a heat shock at 42°C for 60 sec, after which the cells were placed on ice for 2 min. Preheated LB medium (Luria-Berthani sterile medium containing 10% w/v tryptone, 5% w/v yeast extract, 10% w/v NaCl, pH 7.5; 800 µL, 37°C) was added to the transformed cells and the cells grown with shaking (200 rpm) at 37°C for 1 h. Transformed cells (100 uL) were plated out on LB-amp agar plates (LB-broth with 1% w/v agar and 50 µg/mL ampicillin) and grown overnight at 37°C.
For the preparation of heat shock competent cells 5 mL of LB-broth was inoculated with E. coli BLR (DE3) and grown overnight at 37°C in a shaking incubator at 200 rpm. The cells were diluted 1:100 into 250 mL fresh LB-broth and were incubated at 37°C with shaking (200 rpm) until an optical density (OD) at 600 nm of 0.5 was reached. The cells were then cooled on ice for 15 min and subsequently centrifuged at 3000g for 15 min at 4°C. Whilst working on ice, the remaining pellet was gently re-suspended with 50 mL 80 mM cold CaCl2 and left on ice for 40 min. The cells were centrifuged at 3000g for 15 min at 4°C and the pellet was again re-suspended with 5 mL 80 mM cold CaCl2 as well as 2 mL 50% w/v glycerol. Cells were divided into 100 µL aliquots and frozen at -70°C.
Selected single colonies were used to inoculate LB broth (10 mL) containing 50 µg/mL ampicillin (LB-amp) and grown overnight at 37°C. Cultures were diluted 1:100 into LB-amp media and grown to an OD600nm of 0.5. Expression was induced using 200 ng/mL anhydrotetracycline (IBA, Germany) and grown for 4 h at 37°C with shaking. Cells were centrifuged at 2000g for 10 min (Avanti J-26 XP, Beckman Coultier) and the remaining pellets were re-suspended using 15 mL buffer W, consisting of 100 mM Tris-HCl, 150 mM NaCl adjusted to pH 8.0 using HCl. Phenylmethylsulphonyl fluoride (PMSF, 0.1 mM) and 5 mg lysozyme (Sigma Aldrich) was added for every 10 mL re-suspended bacterial cell pellet. The re-suspended pellets were sonicated at 40 kHz using pulsing at output 6, duty cycle 40 with 1 min resting intervals on ice (Sonifier 250, Branson). The lysed bacterial mixture was clarified by centrifugation at 50000g for 50 min at 4°C (Eppendorf).
Summary
Abbreviations
Table of Contents
List of Figures
List of Tables
Acknowledgements
Chapter 1
1.1 Epidemiology of malaria
1.2 Control and treatment strategies
1.3 The significance of vitamin B6
1.4 Hypothesis, research objective and aims
1.5 Outputs
Chapter 2
2.1 Introduction
2.2 Methods
2.3 Results
2.4 Discussion
2.5 Conclusions
Chapter 3
3.1. Introduction .
3.2. Methods
3.3. Results
3.4. Discussion
3.5. Conclusions
Chapter 4
4.1 Introduction
4.2 Methods
4.3 Results
4.4 Discussion
4.5 Conclusions
Chapter 5
References
GET THE COMPLETE PROJECT