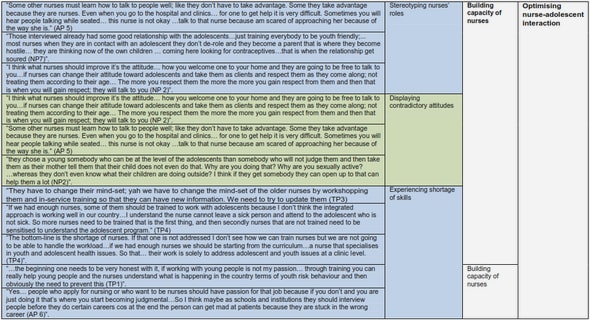
Get Complete Project Material File(s) Now! »
CHAPTER 2 LITERATURE REVIEW
DETERMINATION OF MEAT QUALITY
The properties of meat are determined by several factors spanning from the conception of the animal to the consumption of the meat (Hofmann, 1994). These factors determine the quality of meat as described by indices such as pH, colour, tenderness, flavour, juiciness and nutritive value. In this section, some of the major processes in the evolution of meat quality are reviewed, focusing on how they affect meat quality in general and the quality of chevon specifically.
Myofibre and muscle metabolic types
Muscles are classified into metabolic types on the basis of their predominating myofibre types.There are four myofibre classes which are determined by the metabolic and contractile properties of their constituent myofibres. The three major types are the red (type I or β-red); intermediate (type IIA or α-red) and white (type IIB or α-white) (Brooke and Kaiser, 1970; Ashmore and Doerr, 1971). The fourth class, type IIC exists commonly in neonates and is a transitory link in the formation of types IIA and IIB (Young, 1984; Brandstetter, Picard and Geay, 1998a).Type I myofibres are the smallest in diameter (Rosser, Norris and Nemeth, 1992). They are associated with more blood capillaries, a high lipid, myoglobin, mitochondria and tricarboxylic acid (TCA) cycle enzyme content to suit their high oxidative metabolism (Essén-Gustavsson,Karlström and Lundström, 1992). Strong succinate dehydrogenase (SDH) activity is thus used to identify the myofibres histologically (Brooke and Kaiser, 1970; Ashmore and Doerr, 1971;Swatland, 1981). At the other end, type IIB myofibres are the largest (Rosser et al., 1992). They hold more readily available energy compounds such as creatine phosphate, adenosine triphosphate (ATP) but less glycogen than red muscles (Monin, 1981; Rosser et al., 1992).
Histologically they are distinguished by strong ATPase and lactate dehydrogenase (LDH) activity but weak SDH activity (Brooke and Kaiser, 1970; Ashmore and Doerr, 1971; Swatland,1981). Type IIA myofibres are intermediate in size (Rosser et al., 1992). Their metabolic activity however may either be greater than or intermediate between type I or type IIB depending on the species considered (Ashmore, Tompkins and Doerr, 1972; Monin, 1981).In line with myofibre classification, muscles are classified into red (type I), intermediate (type IIA) and white (type IIB). Red muscles have a high proportion of type I myofibres. They are predominantly postural muscles (e.g. M. trapezius* in the shoulders; M. semimembranosus in the hind leg), with high oxidative capacity to meet the requirements for stamina (Totland and Kryvi,1991; Essén-Gustavsson, 1996). Muscles involved in locomotion (e.g. M. semitendonosus in the hind leg) have a higher glycolytic than oxidative capacity for rapid contraction and so are dominated by the type IIB myofibres. Within individual muscles there is a topographical variation in myofibre type. For example, deeper regions of M. semitendonosus (ST) tend to be darker and more oxidative than outer ST regions (Dreyer, Naudé, Henning and Rossouw, 1977;Hunt and Hedrick, 1977; Totland, Kryvi and Slinde, 1988). In addition, deep type IIB myofibres are more oxidative and have smaller diameters than the superficial ones (Rosser et al., 1992).Muscle metabolic type is also influenced by variations between animals, such as species, breed,sex, age, weight, nutrition and exercise (Essén-Gustavsson, 1996). An example of species differences is the classification of some ovine and bovine muscles (Figure 2.1). The remarkable features of this classification are that, firstly, the type IIA ovine muscles are more glycolytic than the type IIB and more oxidative than the type I. In cattle however, type IIA muscles are intermediate between type IIB and type I in glycolytic and oxidative activities. Secondly, the same muscle may classify differently in different species. For instance whereas the M.semimembranosus (SM) muscle is type IIA in sheep, in cattle it is type IIB (Monin, 1981).Sex and age effects on myofibres were illustrated by Spindler, Mathias and Cramer (1980) who reported a twofold increase in the cross-sectional area of M. biceps femoris (BF) myofibres in steers and heifers ranging from 28 to 392 days old. As the animals grew the myofibre profile increased in type IIB and decreased in type IIA. Such changes in myofibre size and profile were also observed in later works (e.g. Seideman, Crouse and Cross, 1986; Jurie, Robelin, Picard and Geay, 1995; Brandstetter et al. 1998a). They are a result of the general differentiation pathway of type I → type IIA → type IIB (Ashmore et al., 1972) during early stages of muscle hypertrophy Amongst the males, castration effects on muscle fibre type composition are manifested postpubertally. Most reports, such as Dreyer et al. (1977), Young and Bass (1984), Seideman et al.(1986) and Mohan Raj, Moss, McCaughey, McLauchlan and McGaughey and Kennedy (1991) suggest that the proportion of type I myofibres is relatively unaffected by castration while type IIB increase at the expense of type IIA. Brandstetter, Picard and Geay (1998b) however reported
pre-pubertal differences between bulls and steers; that bull calves start showing a tendency to a typical myofibre composition from as early as when they are four months old.Furthermore Brandstetter et al. (1998b) observed that bulls increased in type I and decreased in type IIB myofibres while steers increase in IIB myofibres, but the proportion of IIA myofibres remained unchanged in both sexes. The argument for these changes was that androgens promote an ageing kind of differentiation in myofibres by favouring a shift to type I myofibres (Powers and Florini, 1975). On the other hand, castration delays this re-conversion, and hence the steers had a myofibre type composition that was physiologically less mature than that of bulls of similar age. Despite the differences in the reports on early myofibre type proportions in bulls and steers, most studies agree that androgens promote myofibre hypertrophy, and hence myofibres of steers, particularly types IIA and IIB, tend to be smaller than those of bulls (Dreyer et al., 1977; Young and Bass, 1984; Seideman et al., 1986; Brandstetter et al., 1998b; Dalle Zotte,Verdiglione, Rémignon, Cozzi, Andreoli, Gottardo and Andrighetto, 2000).In a study involving the three sex classes, Young and Foote (1984) suggested that the proportion of type I myofibres is unaffected by the sex of cattle but the proportion of type IIB myofibres of females lay between that of bulls and steers. Conversely, the proportion of type IIA myofibre is higher in female cattle than in steers (Johnston, Moody, Boling and Bradley, 1981; Young and Foote, 1984).Energy restriction leads to a reduction in myofibre size with a strong effect on type IIB atrophy (Yambayamba and Price, 1991; Ward and Stickland, 1993). Conversely, increased dietary energy results in a higher proportion of type IIB and less type IIA myofibres, while high protein diets appear to decrease the proportions of both type IIA and IIB myofibres (Johnston, Stewart, Moody, Boling and Kemp, 1975). Sex effects have been observed in myofibre response to nutrition. Brandstetter et al. (1998b) noted that energy restriction and re-alimentation did not affect the myofibres of steers. Energy restricted bulls however increased oxidative activity but, on re-alimentation, the physiological: chronological myofibre profile was re-established.Yambayamba and Price (1991) observed a similar re-establishment of ‘normal’ myofibre composition and size on re-alimentation of previously restricted heifers.Prolonged physical exertion generally causes age related changes on muscle fibre composition. It increases the proportion of oxidative myofibres, oxidative capacity, capillary density of myofibres, myoglobin content and glycogen storage capacity (Aalhus and Price, 1991). Such changes are expected to result in tough meat because red myofibres have less glycogen, are prone to cold shortening and have thicker z-line that are less susceptible to degradation postmortem (Aalhus, Price, Shand and Hawrysh, 1991). However, Aalhus et al. (1991) reported tender SM from exercised compared to none exercised sheep. This was alluded to the increase in the myofibrillar-to-collagen protein ratio with exercise than to changes in myofibre type composition .
Implications of myofibre composition on sampling for meat quality evaluation
Variations in myofibre composition within muscle in addition to the inter-muscle differences suggest that representative sampling procedures should be employed for meat quality evaluations. In the majority of meat science studies, the M. longissimus thoracis et lumborum (LTL) is used as the standard muscle for the evaluations. However, in goats this muscle is too small to obtain enough samples for all the standard procedures and hence other muscles such as the SM have also been used along with the LTL muscle (Babiker, El Khidir and Shafie, 1990;Schönfeldt et al., 1993a and b; Swan, Esguerra, and Farouk, 1998). There has been a suggestion that the two muscles are of similar type in cattle and sheep (Pethick, Cummins, Gardner, Jacobs,
Knee, McDowell, McIntyre, Tudor, Walker and Warner, 2000) but there is no known classification of these muscles in goats. An understanding of the myofibre profile of the both the LTL and SM of goats would therefore be beneficial in making inferences on quality attributes observed on these muscles.
Abstract
Declaration
Summary
Acknowledgements
List of acronyms
List of Tables
List of Figures
1 INTRODUCTION
1.1 Project theme
1.2 Project title
1.3 Aims
1.4 Motivation
2 LITERATURE REVIEW
2.1 Determination of meat quality
2.1.1 Myofibre and muscle metabolic types
2.1.1.1 Implications of myofibre composition on sampling for meat
quality evaluation
2.1.2 Conversion of muscle to meat
2.1.2.1 Development of rigor mortis
2.1.2.2 Post-mortem glycolysis
2.1.2.3 Rate of post-mortem glycolysis
2.1.2.4 Extent of post-mortem glycolysis
2.1.2.5 Post-mortem tenderisation
2.1.2.5.1 The calpains
2.1.2.5.2 Calpastatin
2.1.2.5.3 Factors influencing concentration of calpains
2.2 Meat quality factors
2.2.1 Meat colour
2.2.1.1 The colour of chevon
2.2.2 Water in meat
2.2.2.1 Water losses in chevon
2.2.3 Fat in meat
2.2.3.1 Fat in chevon
2.2.4 Meat juiciness
2.2.4.1 Juiciness of chevon
2.2.5 Meat flavour and aroma
2.2.5.1 Flavour and aroma of chevon
2.2.6 Meat tenderness
2.2.6.1 Collagen and its contribution to meat tenderness
2.2.6.2 Myofibrillar contribution to tenderness
2.2.6.2.1 Pre-slaughter factors
2.2.6.2.2 Post-slaughter factors
2.2.6.3 Tenderness of chevon
2.2.7 Factors of production quality
2.2.7.1 Effect of nutritional history.
2.2.7.2 Effect of physical exercise
2.2.7.3 Effect of peri-mortem treatment
2.2.7.4 Effect of post-slaughter handling
2.2.8 Implications of smallholder production systems on chevon quality
2.3 Sensory evaluation of meat quality
2.4 Summary
3 MATERIALS AND METHODS
3.1 The experimental goats
3.2 Sampling and sample storage
3.3 Carcass measurements
3.4 Histological and histochemical analysis
3.4.1 Sarcomere length
3.4.2 Myofibrillar fragment length
3.4.3 Myofibre typing
3.5 Physical meat characteristics
3.5.1 Colour, cooking losses and shear force
3.6 Biochemical analyses
3.6.1 Glycolytic potential, ATP and creatine phosphate
3.6.2 Calpastatin
3.6.3 Fatty acids
3.6.4 Amino acids
3.6.5 Crude nitrogen and crude protein
3.7 Sensory evaluation
3.7.1 Preparation of samples for sensory evaluation
3.7.2 Sensory panels and sensory evaluation
3.8 Statistical analysis
3.8.1 Live animal carcass and meat quality characteristics
3.8.1.1 Live animal and carcass characteristics
3.8.1.2 Meat quality of chevon
3.8.1.3 Effects of electrical stimulation on chevon quality
3.8.2 Fatty acid and amino acid composition
3.8.3 Sensory evaluation
4 LIVE ANIMAL AND CARCASS CHARACTERISTICS OF
SOUTH AFRICAN INDIGENOUS GOATS
4.1 Introduction
4.2 Results
4.2.1 Live animal and carcass characteristics
4.2.1.1 Effect of sex on live animal and carcass characteristics
4.2.1.2 Effect of age on live animal and carcass characteristics
4.2.1.3 Effect of pre-slaughter conditioning on live animal and carcass characteristics
4.1.2.4 Interaction effects of sex, age and pre-slaughter conditioning on live animal and carcass characteristics
4.2.2 Carcass composition
4.2.2.1 Effect of sex on carcass composition
4.2.2.2 Effect of age on carcass composition
4.2.2.3 Effect of pre-slaughter conditioning on carcass composition
4.2.2.4 Interaction effects of sex, age and pre-slaughter conditioning on carcass composition
4.3 Discussion
4.3.1 Live animal and carcass characteristics
4.3.2 Joint and tissue composition of the carcasses
4.4 Summary
5 MEAT QUALITY CHARACTERISTICS OF CHEVON
5.1 Introduction
5.2 Results
5.3 Discussion
5.4 Summary
6 THE EFFECT OF ELECTRICAL STIMULATION ON
CHEVON QUALITY
6.1 Introduction
6.2 Results
6.3 Discussion
6.4 Summary
7 FATTY ACID AND AMINO ACID COMPOSITION OF
CHEVON
7.1 Introduction
7.2 Results
7.3 Discussion
7.4 Summary
8 ACCEPTABILITY OF CHEVON TO SOUTH AFRICAN
CONSUMERS
8.1 Introduction
8.2 Results
8.3 Discussion
8.4 Summary
9 INTEGRATION, CONCLUSIONS & RECOMMENDATIONS
9.1 Integration
9.2 Conclusions
9.3 Implication of findings
LIST OF REFERENCES
GET THE COMPLETE PROJECT
Meat characteristics and acceptability of chevon from South African indigenous goats